
Pannell Lab Overview
Current projects in the group are addressing questions concerning transitions between sexual systems and their implications for mating, resource allocation, demography and the evolution of the genome and transcriptome, especially sex chromosomes. Some of these are summarised under the project descriptions below. These projects use a combination of theory, wet-lab work, bioinformatics, and fieldwork. See our recent publications for further details of results from these and other projects.
Transitions between combined and separate sexes in Mercurialis annua
Much of our research aims to explain and understand transitions between combined and separate sexes, and the evolution of sex chromosomes, in the European annual plant Mercurialis annua, which displays dioecy (fully separate sexes), monoecy (self-fertile functional hermaphroditism) and androdioecy (the coexistence of males and hermaphrodites) and male sterility. M. annua has an assembled and annotated genome and shows many of the hallmarks of Y-chromosome degeneration. Interestingly, male-function loci regulating pollen production are not located on the Y chromosome, but the secondary sexual dimorphism influencing inflorescence architecture is Y-linked and has introgressed among related species, likely due to strong frequency-dependent selection.
Related projects are examining phenotypic changes in sex allocation, changes in gene expression, and the underlying genomic basis of these changes in experimental populations of M. annua in which mate availability has been manipulated.

Male (A), female (B) and monoecious (C) individuals of Mercurialis annua. Photos: J. Baker.
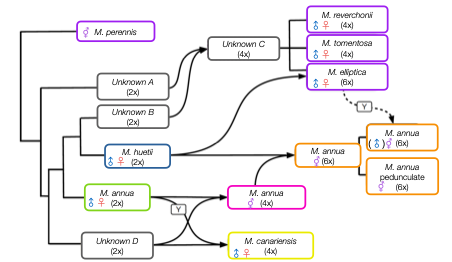
Phylogenetic reconstruction of the genus Mercurialis on the basis of variation at a large number of loci at autosomal and sex-linked loci reveals a complex history of hybridization and genome duplication, transitions between sexual systems and the introgression of a Y chromosome into androdioecious populations of M. annua (in which males co-occur with hermaphrodites) from a more distantly related (perennial) species. Image: Jörn Gerchen and Paris Veltsos.
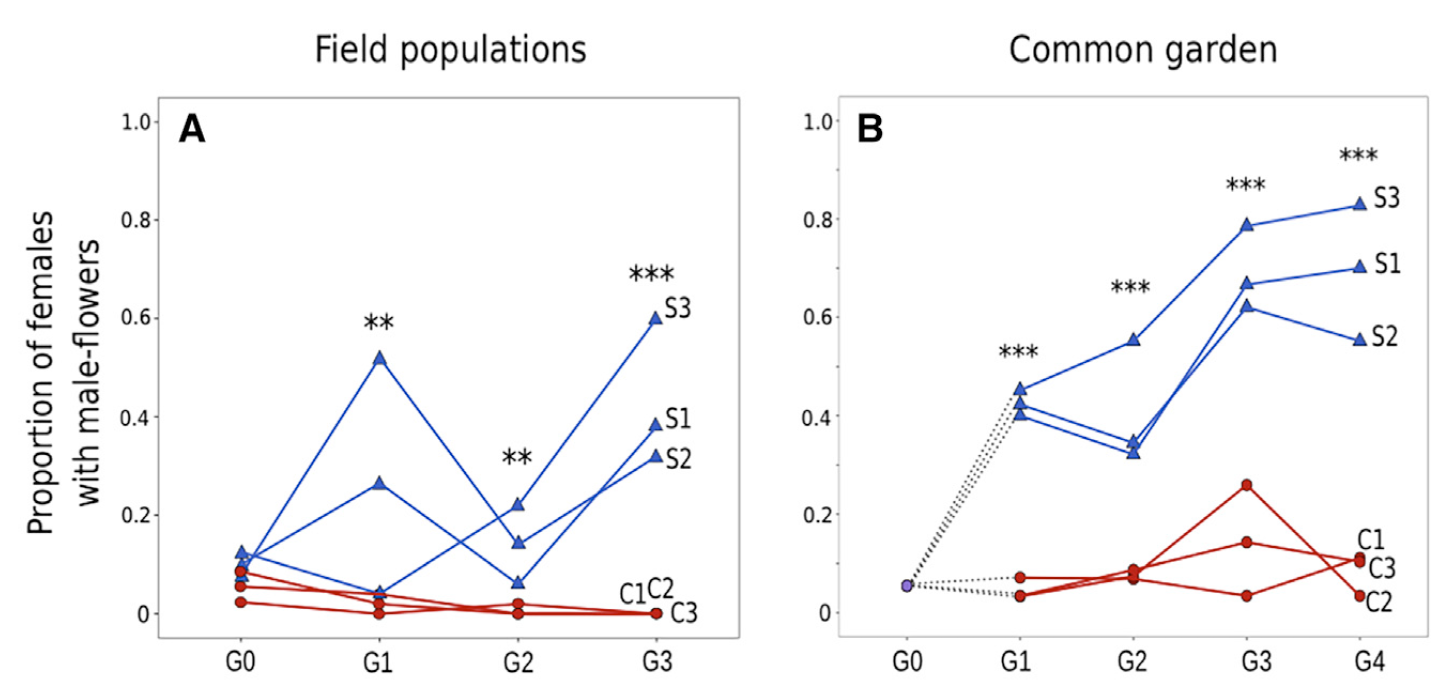
In a key ongoing experiment, the removal of males from dioecious populations has led to the evolution over several generations of substantial male expression by females and thus a rapid transition from dioecy to monoecy. The figure shows the rapid evolution of sex allocation in ‘leaky’ females of the dioecious plant Mercurialis annua after the experimental removal of males from three populations (blue lines) compared with those in populations in which males were maintained (red lines). Patterns measured in the field experimental populations were similar to those measured in a common garden in which all genotypes from all generations (G0 – G4) were grown together under the same conditions. Females evolving in the absence of males dramatically increased their male allocation. Graphs from Cossard et al. (2020, Current Biology; see reference list).
Evolution of sexual dimorphism
Although plants do not usually show morphological and other differences between males and females as strikingly as do animals, sexual dimorphism in some plants can be strong. We are interested in the ecological contexts in which different strategies are selected in males and females of dioecious plants, and the way that natural and sexual selection modify their genomes and the transcriptomes. Here, we are using as models both the striking variation among sexual systems in M. annua as well as the variation among species in terms of sexual dimorphism in the South African genus Leucadendron. We recently showed that genes with sex-biased expression among 10 species of the genus sampled widely across the phylogeny have elevated rates of expression evolution. While this might suggest a role of sexual selection in driving expression evolution, our analysis indicates that in fact the sex-biased genes had been evolving more rapidly than average before they had become sex-biased, suggesting that sex-biased genes might be drawn from a class of genes that are less constrained in their expression patterns.

Typical differences in morphology displayed in the South African genus Leucadendron, in which males (left) are more often highly branched, with smaller leaves, than females (right). Photo: John Pannell
Evolution and ecology of sex allocation
Although hermaphroditic plants invest their resources in both male and female functions, they may benefit more from one sex than the other under different conditions. We are studying the very broad variation in sex allocation in the Alpine perennial herb Anemone alpina in an attempt to understand how plants make their allocation decisions. Our work has found that floral traits affect fitness in ways that depend on sex allocation, though the attraction and manipulation of pollinators, but also of herbivores and their parasitoids. We are currently using genetic markers and multi-year data on fitness components estimated for tagged individuals to infer the relationship between allocation, morphological architecture and male and female fitness.

Variation in sex allocation in the Alpine perennial plant Anemone alpina. A: female flower; B: hermaphrodite flower; C: Male flower; D: individual with multiple flowers varying in their sex allocation. Populations of A. alpina comprise individuals that differ from expressing only male function to those expressing both male and female functions. The between-season variation in allocation for an individual depends on its resource status, which is influenced by its size, its sex allocation in the previous season, and whether it has been subject to herbivory. Photos: Kaihsiu Chen.
Inferring paths of sexual-system evolution in the context of genome duplication
We are interested in the affects that demographic processes such as metapopulation dynamics, population-size fluctuations, and range expansions have of the evolution of the mating system, sex allocation and the adaptive potential of populations. Our research here uses both demographic and population-genetic simulations, as well as bioinformatic analysis of NGS gene-capture data. We have been using Approximate Bayesian Computation to understand the order of key that demographic events, hybridization and whole-genome duplication have played in the diversification of plant lineages.
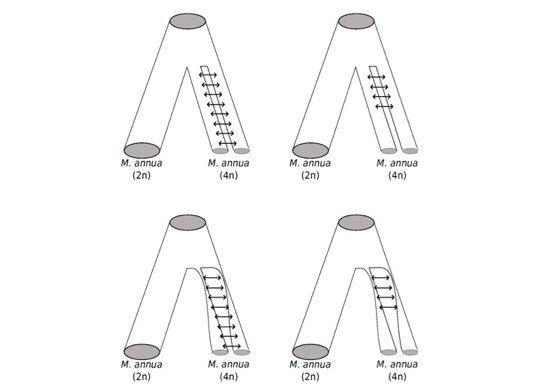
Diagrams depicting four of the many possible scenarios underlying genome duplication via autotetraploidy and subsequent diploidisation of the genome (the cessation of gene flow between homoleogous genomes). Tube width reflects population size. Approximate Bayesian Computation is being used to discriminate among these and other scenarios. Image: Camille Roux.
We have also used transcriptome data to infer the parental origin of the different homeologous genomes of the allopolyploid dioecious species Mercurialis canariensis and the fate of the different genomes in terms of sex-biased gene expression.
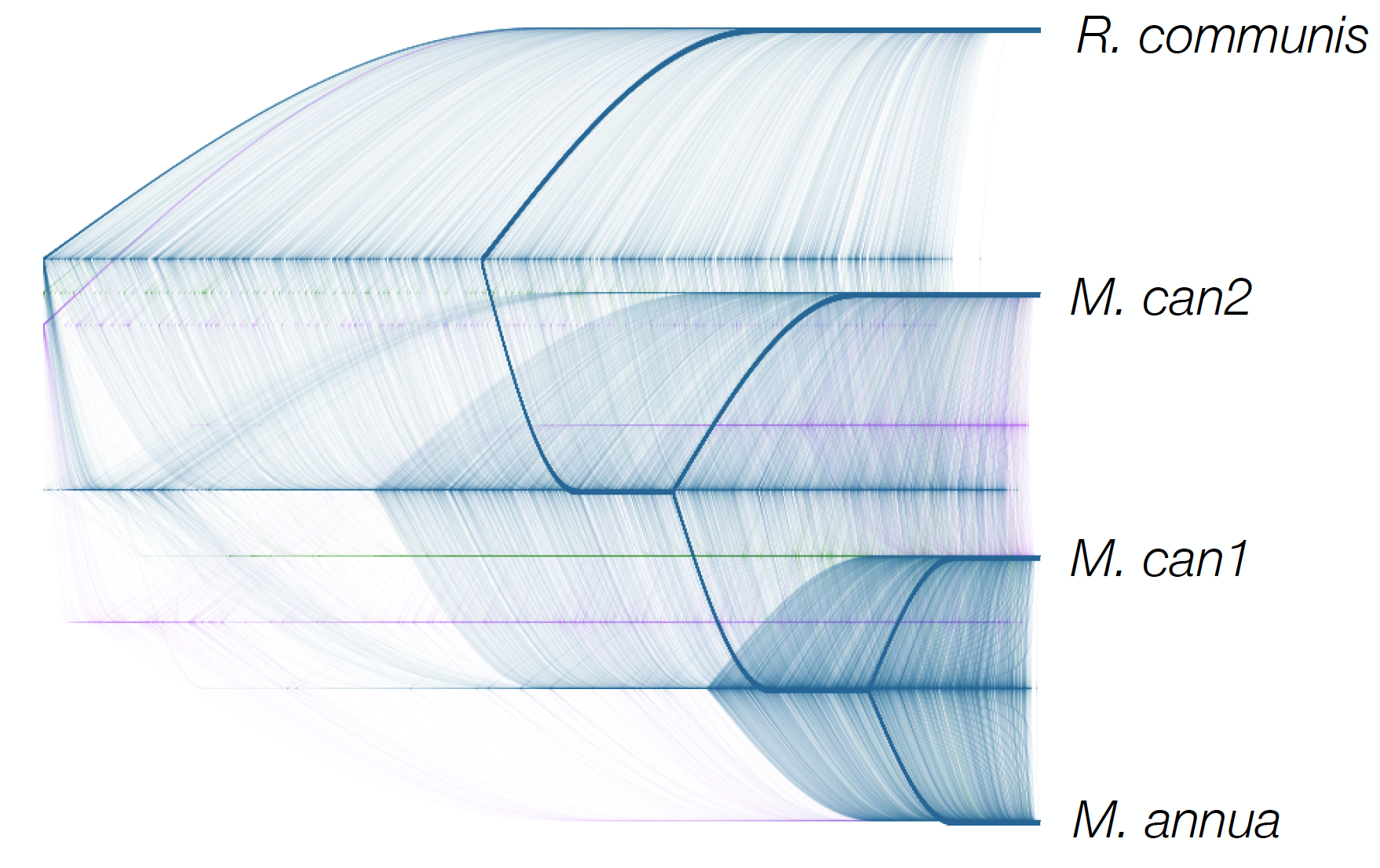
The variation in the topology of 3135 gene trees for in the inferred phylogeny linking Ricinus communis, Mercurialis annua, and the two subgenomes of M. canariensis. The graph indicates that genes in M. can1 tend to be more closely related to M. annua (and were derived from a more recent ancestor of the two lineages) than those in M. can2 (which have a more ancient common ancestor). The Y chromosome of M. canariensis is derived from M. annua. Image: Melissa Toups.
Evolutionary ecology of flower and inflorescence strategies under wind pollination
We are interested in the demographic implications of both sexual dimorphism and pollen limitation, particularly in wind-pollinated species in which mating interactions can be modelled and measured as a function of density. Our work in this domain employs comparative meta-analysis, mathematical modelling and manipulative experiments. We are particularly working on the demographic implications of selection on sex-allocation and inflorescence traits that are thought to be sexually antagonistic.

Experimental arrays of wind-pollinated Mercurialis annua, in which distance from a pollen source regulates the intensity of pollen competition. Photo: A-M. Labouche.
Social evolution in plants
It is becoming increasingly appreciated that not only animals but also plants may have a social life, and that social interactions among plants that differ in their relatedness with one another can bring about the evolution of both selfish and altruistic behaviour. Working with the attractive insect-pollinated plant brassica Moricandia moricandioides as a model, we found that groups of related individuals were more likely to invest towards attracting pollinators to the whole group (a sign of altruistic behaviour) than individuals growing with unrelated neighbours.

Inflorescence of the plant Moricandia moricandioides, used as a model in experiments on the social evolution of plants. Photo: Rubén Torices.
Evolution of senescence in plants
Do all plants age? Some plants, such as the spectacular Echium wildpretii, which grows on trhe slopes of Teide in Tenerife, are monocarpic: they flower once, and then die. Others can live for thousands of years, with no evidence of physiological decline. In collaboration with the Mullon group in Lausanne, we are currently working on theoretical models to understand when plants should senesce, and how plants might limit their ageing process.

The Canary Islands endemic Echium wildpretii, a monocarpic perennial that flowers once, after several years of growth, and then dies. Photos: John Pannell.
Population ecology and genetics of range expansions
Range expansions often involve repeated genetic bottlenecks that are predicted to allow deleterious mutations to reach high frequencies of fix in populations more than they would in large non-expanding population. The bottlenecks also reduce the effective population size of the range edge populations such as to reduce the efficacy of positive selection, so that we expect fewer ‘selective sweeps’ in the genome, where positively selected mutations have been taken to high frequency by natural selection.
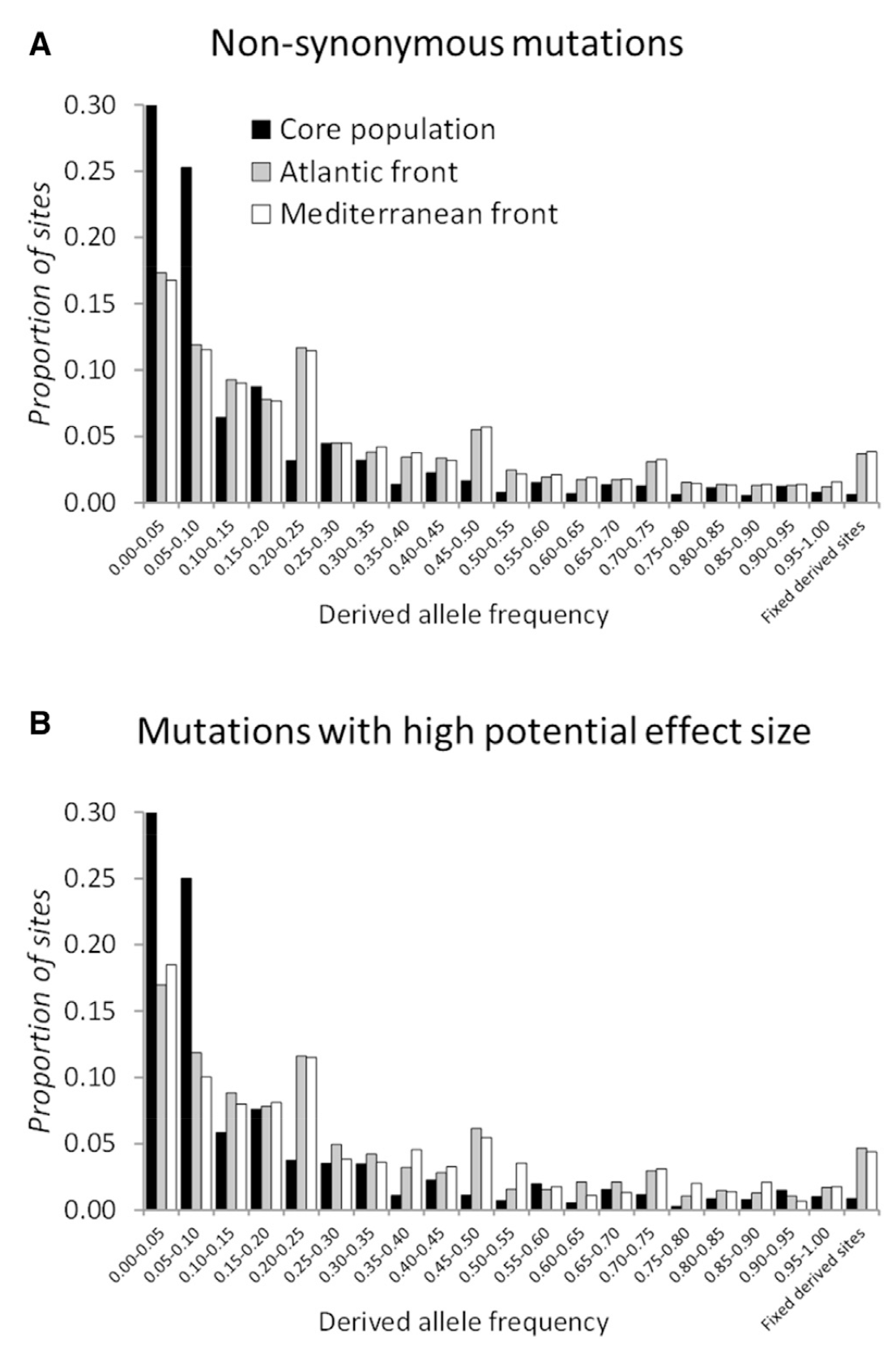
This figure shows the site frequency distributions for derived mutations of likely (A) small and (b) larger fitness effects on phenotypes of the dioecious annual colonising plant Mercurialis annua. Populations were sampled in the eastern Mediterranean ‘core’ of gthe species distribution, prior to range expansion, and two sets of range-edge populations (Mediterranean front and Atlantic front populations). The graphs indicate an increase in likely deleterious mutations in the range-edge populations. Figure from Gonzalez-Martinez et al. (2017, Current Biology; see references).
Group leader
Post-Docs
Graduate Students
Staff
- Aline Revel
Former Group Members
- Dr. Regina Berjano
- Dr. Richard J.A. Buggs
- Ruben Torices Blanco
- Dr. Gill Campbell
- Dr. Guillaume Cossard
- Yves Cuenot
- Dr. Esther Dale
- Dr. Marcel Dorken
- Jonathan El Assad
- Dr. Sarah M. Eppley
- Dr. Jörn Gerchen
- Dr. Santiago Gonzalez-Martinez
- Dr. Alok Gupta
- Dr. Mark Harris
- Dr. Elze Hesse
- Dr. Dan Jeffries
- Francisco Javier Jimenez
- Dr. Grazyna Korbecka
- Dr. Anne-Marie Labouche
- Xinji Li
- Dr. Juan-Carlos Lopez-Almansa
- Dr. Wen-Juan Ma
- Ana Paula Machado
- Thomas Martignier
- Dr. Jamie Moore
- Dr. Darren Obbard
- Dr. Benoit Pujol
- Dr. Kate Ridout
- Isabel Rodriguez
- Dr. Rebecca Ross
- Dr. Camille Roux
- Luka Rubinjoni
- Nicolas Ruch
- John Russell
- Solenn Sarton-Lohéac
- Dr. Mathias Scharmann
- Lucia De Soto Suarez
- Aline Revel
- Dr. James Gordon Rodger
- Dr. Paul Rymer
- Dr. Julia Sanchez-Vilas
- Dr. Luis Santos del Blanco
- Maria Talavera Solis
- Dr. Thomasz Suchan
- Nora Szijarto
- Dr. Jeanne Tonnabel
- Dr. Melissa Toups
- Simon Vogel
- Dr. Paris Veltsos
- Dr. Nora Villamil Buenrostro
- Dr. Marie Voillemot
- Neil White
- Dr. Melissa Wong
- Dr. Shurong Zhou
